Locomotion
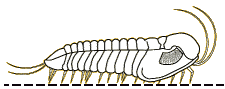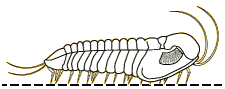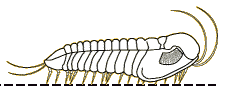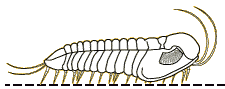
For the most part, trilobites move in a way you would expect them to move. Trilobites were mostly benthic creatures, meaning that they were crawling across the seafloor (Benton & Harper, 2009). However, some trilobites were pelagic, or freely swimming in the water column (Benton & Harper, 2009). The benthic trilobites had very flat, spiked bodies, which were not suited for swimming and fast mobility but rather for defensive purposes and moving across loose muddy sediment (Benton & Harper, 2009). The benthic species typically moved straight forward much like a modern pillbug (Clarkson, 2013). However, from looking at fossilized trilobite footprints, we can see that they had other directions of locomotion. Some sets of fossils had their footprints closer together on one side and more separate on the other side (Clarkson, 2013). This implies that trilobites were able to move obliquely (that is, diagonally), or side to side, like modern day crabs do (Clarkson, 2013). Their appendages move in a metachronal rhythm, meaning that limbs move forward slightly and pushing backwards, helping to support the body, and moving the trilobite forward (Clarkson, 2013). This process is repeated by the next legs in front in a wave-like motion, starting from the pygidium all the way up to the cephalon (Clarkson, 2013). The pelagic species were more streamlined, with a flexible exoskeleton, a long slender body, and large eyes, very similar to modern shrimp (Benton & Harper, 2009). There are a wide variety of trilobite morphologies, which were related to the trilobites’ various lifestyles (Benton & Harper, 2009). Trace fossils from burrows and tracks suggest that trilobites could burrow into sediments to feed or avoid predators (Benton and Harper, 2009).

Picture of the different types of locomotion of trilobites (University of Maryland, 2016)
Body Plan
The name “Trilobite” comes from the Latin word three-lobed. This describes the common body plan that most species within the Trilobite phylum which is indicative of how the exoskeleton is divided into a central axis with two lobes, one on the left and the right (Bell, 2013). The left and right lobes are called left and right pleural lobes respectively (Bell, 2013). Also, their bodies are divided into three sections called tagmata from front to back. The first section from the head is called a head shield, specifically the cephalon, a body that is separated into segments called the thorax and a tail shield which consists of multiple fused segments called the pygidium (Benton and Harper, 2009). From this common body plan, there is a great amount of diversity in species belonging to Trilobites. The earliest forms of this body plan included a small pygidium, well developed eye ridges flattened body form and several thoracic segments. The diversification of body plans either had a mixture of an elongated form as seen in Crotalocephalina sp., a widened body seen in Harpillaenus sp., increased thoracic segments in Balcoracania sp., and/or reduced segments in Thoracocare sp. (Gon, 2013). Morphological diversity increased from the Cambrian period and peak diversification of Trilobites occurred in the Ordovician.


Figure of a line drawing of a common trilobite plan showing the three main parts of the body, cephalon, thorax, and pygidium (Gon, 2013).
Figure of a line drawing of a common trilobite body plan showing the "three lobes"; left pleural lobe, axial pleural lobe and right pleural lobe (Gon, 2013).
Trilobites vary in immensely in size and in morphology. The smallest Trilobite fossils that have been found can be as long as 1-2 mm such as the Acanthopleurella stipullae. The largest Trilobite fossil was found in Manitoba, Canada which indicates some species grew to as large as the 720mm such as the Isotelus rex (Whittington and Kaesler, 1997). Some species have been found to show special features such well-developed eye systems, spines, smoothened body appearance, and a loss of eyes or reduced eye systems (Benton and Harper, 2009). A smoothened body appearance was thought to be due to the burrowing behaviour of some trilobites (Gon, 2013). In several orders of Trilobites such as the Asaphida or Agnostida, are characterized by a smoothened appearance most likely due to a burrowing lifestyle. Another reason for this smoothened appearance might have been due to making the body more streamline to move better in the pelagic waters (Gon, 2013). Spines are commonly thought to be primarily a defense mechanism and diversity in the number of spines is seen in a number of trilobites. They are also thought to be an adaptive structure for stabilization on uneven or loose surfaces, or flotation/ stabilization structures for certain trilobites (Gon, 2013).


Picture showing the spinosity in Leonaspis sp. (Gon, 2013).
Picture of Isotelus rex, one of the largest trilobites
Anatomy of Trilobites can be further divided into internal, dorsal (back), and ventral (underside) structures. Internal anatomy consisted of basically a mouth connected to the stomach and a straight long canal to the anus where waste was excreted (Gon, 2013). It also had gastric caecae which were thought to be similar function to a liver (Gon, 2013). Dorsal or back structures had a glabella – middle portion of the cephalon (head) typically lobed and convex, combined with furrows (grooves) and sutures (breaks in skeleton) which typically separate during molting (Gon, 2013). Based on these furrows and sutures Trilobites can be classified and put into families and classes. Dorsal sutures can be divided into 5 types, absent – lacking facial sutures (e.g. Olenellina), proparian – facial suture ends in front of the genal angle, gonatoparian – facial suture ends at the tip of the genal angle, opisthoparian, faicla suture ends at the posterior margin, and hypoparian – derivative of opisthoparion suture, which lost facial sutures. Hypoparian was thought to be a secondary loss or reversion of the dorsal suture trait (Frank, 1949). The ventral sutures are connective sutures which divide the doublure (an extension of the dorsal exoskeleton which produces a fringe) (Gon, 2013).


Figure of a line drawing of the general internal anatomy of a trilobite (Gon, 2013).
Picture of Selenocoryphe platyura. note the middle of the fossil showing the alimentary canal running along the whole midline of the body (Gon, 2013).
Special Features- Eyes
Underside structures consisted of a hypostome which is the calcified mouth of the trilobite. Limbs are also present in the underside of the trilobite which are attached to the axial midline and at the tips of the limbs were jagged toothlike structures that helped the trilobite pass food from the ground to the mouth (Fortey and Owens, 1999). Some species also had antennae which acted like sensory organs (Gon, 2013).

Figure of a line drawing of the ventral (underside) of Olenoides serratus. Note the large calcified hypostome along with the limbs attached to the axial midline (Gon, 2013).
Eye systems in Trilobites are some of the most studied topics about this organism. They were some of the first organisms to develop such a complex visual system on earth at the time. The majority of Trilobites have compound eyes. Compound eyes are eyes that are made up of many lenses which occupy the outer sides of the cephalon (head) of Trilobites (Thomas, 2005). The advantages of having compound eyes are that they are very sensitive to motion which may have aided in predator detection or stalking prey (Gon, 2013). It is also noted that they had rigid crystalline lenses that formed a doublet structure that allowed Trilobites to have an astonishing depth of field that allowed them to see both near and far and minimal distortion of the image (Gon, 2013).

Picture of a trilobite characterized by compound eyes. These eyes have many lenses that give trilobites 360 degree vision (Gon, 2013).
There are three types of Trilobite eyes that have been found in the fossil record; holochroal, schizochroal, and abathochroal (Thomas, 2005). It is thought that abathochroal and schizochroal types have evolved from holochroal eyes (Thomas, 2005). The main difference between these eye types is that holochroal type is large in lenses compared to abathochroal and schizochroal types which have fewer lenses, but larger in size and are separated from one another (Thomas, 2005). Holochroal eyes are characterized by having small, closely spaced lenses covered by a thin cornea. They are usually hexagonal in shape and have many lenses up to 15 000 in each eye! (Thomas, 2005). Abathochroal eyes are characterized by having small but separated lenses with thin scleral walls (white of the eye) and one cornea per lens. (Gon, 2013). Schizochroal eyes are characterized by having larger, thicker lenses that are separated by deep scleral walls (Thomas, 2005). Holochroal eyes are found in nearly all of the Orders of Trilobites, while schizochroal eyes are found only in the Phacopine suborder and abathochroal eyes only found within the Cambrian Eodiscina time period (Thomas, 2005). It has been thought that the schizochroal eye types evolved from holochroal as a type of post-displacement paedomorphosis. Paedomorphosis is the retention of juvenile (young) characteristics into adulthood. Post-displacement paedomorphosis is the delayed growth of certain structures relative to others. With respect to Trilobites, the eyes of immature holochroal Cambrian Trilobites were very similar to miniature schizochroal eyes. In Phacopida, these immature eyes were retained into adulthood and gave rise to schizochroal eye types from holochroal eyes (Thomas, 2005).



Pictures representing the three types of eyes in trilobites. Holochroal (left), Schizochroal (middle), and Abathochroal (right) (Gon, 2013)
The degree of eye variation within the trilobite body plan was extremely diverse. Due to the abundance of Trilobites found in many different ecosystems, variations in eye structures were evident. Many early trilobites had crescent like eyes which resulted in a wide degree of vision; some Trilobites were known to have 360⁰ vision! (Benton and Harper, 2009). Other eye variations included large holochroal eyes which were so large they took up most of the space in the space on the cephalon in organisms such as the Opipeuterella sp (Thomas, 2005). Another variation in Trilobite eyes was the appearance of stalked eyes. These were projections from the cephalon which allowed the eyes to be raised above the body which would help to observe the surroundings for danger or look above rocks or algae in the benthic layer of the ocean (Gon, 2013). An example of a Trilobite with stalked eyes is Neoasaphus sp. (Gon, 2013). A large number of species lived on the bottom layer of the ocean (benthic). This area had very little photic (light) activity so some of the species that lived in this environment might have gradually lost their eyes without suffering an adaptive disadvantage because they were not needed (Thomas, 2005). These Trilobites with reduced eyes or total loss of eyes are called atheloptic (Gon, 2013). Species that were athelopic were Agnostus sp., and the Trimerus sp. (Thomas, 2005).



Picture of stalked eyes of Neoasaphus sp. (Gon, 2013).
Picture of large holochroal eyes of Opipeuteurella sp. (Gon, 2013).
Picture of eyeless Agnostus sp. (Gon, 2013).
Special Features - Protection
Most Trilobites were capable of rolling up into a defensive ball as a means of protection against predators (Gon, 2013). This was done due to the segmentation of the thoracic segments which allowed them to bring the cephalon (head) and the pygidium (tail) together which protected the antennae, limbs and soft underside. While curled up in a defensive stance, they were able to protect themselves until the conditions were safe again and went on their way. Certain species had spines protruding from the exoskeleton to aide in further protection and act as deterrents from being swallowed whole by larger predators (Ortega-Hernandez et al., 2013). Trilobites achieved enrollment by contracting internal muscles thereby bending the flexible shell between the thoracic segments so that the head and tail were brought together. A special feature of some Trilobites is that some such as the Phacopina developed specialized morphological features that enabled them to interlock tightly to form coaptative structures (Gon, 2013).
There are 4 types of enrollment; first is called spaeroidal enrollment when all of the thoracic segments participate in curling of the body. This is the most common type of protection for Trilobites. The other types of enrollment is called the double enrollment characterized by the tail and lower thoracic segments hidden under the front of the cephalon. This is seen in primitive Cambrian trilobites such as Ellipsocephalus sp. (Gon, 2013). Discoidal enrollment also known as basket and lid enrollment is the flexing of the anterior regions of the thorax forming a basket-like structure and the rest of the tail and body covering the flexed thorax like a lid. This is seen in Harpetids and Trinucleioid Trilobites (Ortega-Hernandez et al., 2013). The last type of enrollment is incomplete enrollment where the thoracic segments do not completely close which creates a gap. This is seen in Trilobites with spines as the gap creates a space for the spines to project from. This is seen in spinous Trilobites such as the Olenelloids (Ortega-Hernandez et al., 2013).

Picture of Nyterops sp. undergoing sphaeroidal enrollment (Gon, 2013).

Picture of a harpetid undergoing discoidal enrollment (Gon, 2013).

Picture of Ellipsocephalus sp. undergoing double enrollment (Gon, 2013).